
MODULAÇÃO RADIOINDUZIDA DE RECEPTORES DE CÉLULAS
MONONUCLEARES DO SANGUE PERIFÉRICO EM MODELO ANIMAL
JM Toledo 1, SL Siqueira 2, TPR Campos 1, PL Falcão 1
1 Universidade Federal de Minas Gerais
Programa de Ciências e Técnicas Nucleares
Av. Antonio Carlos, 6627, Bloco 4, S. 2285, Minas Gerais
2 Universidade Federal de Ouro Preto
Departmento de Cirurgia, Ginecologia, Obstetricia e Propedeutica,
Escola de Medicina, Ouro Preto, Minas Gerais.
maio de 2018
Resumo
O presente estudo investiga parâmetros fenotípicos celulares, utilizando marcadores de estimulação e imunosupressão celular em modelo animal. A estratégia experimental consistiu na avaliação de parâmetros fenotípicos de células mononucleares expostas à radiação ionizante, avaliados por citometria de fluxo e comparados pré e pós-cinética de tempo. Foram agrupados seis cobaias irradiados com doses de 2Gy, fracionadas, totalizando 4 Gy e 6 Gy. Amostras do sangue periférico foram coletadas antes (controle) e após as irradiações, nos períodos de 2h, 18h e 30 dias. As células do sangue periférico (PBMC) foram separadas e submetidas às marcações com os anticorpos monoclonais anti-CD5, anti-CD4, anti-IgM, anti-CD14, anti-MHCII, anti-CTLA-4 (CD154) e anti-IL-2R para avaliação das células T circulantes, células B, linhagem monocítica/macrofágica e parâmetros de estimulação, expansão clonal e regulação da expansão clonal, respectivamente. As PBMCs foram marcadas com os anticorpos monoclonais anti-receptores com fluoresceína e ficoeritrina. Os dados foram expressos pelo número absoluto de receptores para os anticorpos de interesse na superfície celular, antes e após a irradiação de cada animal, como média dos percentuais de receptores dos grupos avaliados nos tempos zero (T0), 2h (T2), 18h (T18) e 30 dias após a irradiação (T30), obtidos através do canal médio de fluorescência (CMF). Os achados demonstraram uma diminuição significativa das células T circulantes CD5 e da subpopulação CD4, após a irradiação, bem como a diminuição dos receptores de estimulação e interação celular MHCII e de IL-2R que confere o “start” para a expansão clonal. A diminuição da expressão desses receptores foi acompanhada do aumento da expressão de CTLA-4, que parece ter exercido papel autócrino, regulando a expressão de IL-2R de linfócitos T, bem como de outras populações adjacentes. O papel da linhagem monocítica/macrofágicas, investigado nesse modelo, parece ser controverso, uma vez que a expressão do receptor para CD14 não apresentou uma alteração pontual, sugerindo que sua função de célula apresentadora de antígenos não foi bem definido no modelo investigado, possivelmente proque os danos ocorridos na membrana das células e no núcleo parecem ser bem mais rápidos do que a própria mobilização intracelular. Os dados nos permitiram inferir sobre a importância do conhecimento da resposta imune individualizada, através da investigação do padrão fenotípico das linhagens celulares circulantes sistêmicas.
Palavras - chaves: citometria de fluxo, receptor celular, radiação Co-60
Introdução
As radiações ionizantes promovem alterações biológicas funcionais do organismo humano podendo levar a apoptose ou necrose celular. Essas modificações dependem basicamente do nível da dose absorvida no tecido, da taxa de dose, do tipo de exposição se crônica ou aguda, bem como da área corporal e do tempo de exposição. Quanto maiores as taxas de dose, a dose absorvida e o tempo de exposição, maior será a probabilidade de dano, de mutações precursoras oncogênicas e de morte celular, com prejuízo do metabolismo biológico e com o comprometimento de órgãos e/ou tecidos e de suas respectivas funções orgânicas. (PEREZ, 1998; FRANKS, 1990)
O quadro sanguíneo tem sua funcionalidade alterada após exposição de 0,25 Gy de corpo inteiro. Todas as células do sangue podem sofrer supressão numérica em altos níveis de dose, obedecendo, no entanto, ao seus respectivos graus de radiosensibilidade. Em geral, os primeiros elementos sanguíneos que sofrem supressão em poucas horas são os linfócitos, os granulócitos em alguns dias, as plaquetas e os eritrócitos em algumas semanas. Antes da linfocitopenia há hiperleucocitose passageira dosneutrófilos, com redução dos eusinófilos e, logo a seguir, dos linfócitos. (DEVITA, 1982; STEWART e NICHOLSON, 2000)
Os linfonodos e as formações linfáticas do organismo são altamente radiosensíveis. Observam-se as primeiras alterações a partir de uma dose de 0,25 a 0,50Gy; entretanto, as linhagens são totalmente suprimidas após doses de 4 a 6Gy. Em geral, uma semana após a exposição à radiação ionizante in vivo, inicia-se a fase de regeneração do sistema linfático. Usualmente a regeneração total das linhagens de células brancas ocorre 4 a 6 semanas após as irradiações. (STEWART e NICHOLSON, 2000)
Deve-se ressaltar que as radiações ionizantes podem trazer benefícios para o controle de sítios clonogênicos, bem como efeitos deletérios no tecido sadio. É de extremo interesse ampliar o conhecimento sobre os métodos que nos permitem visualizar e compreender mais claramente o papel dos componentes celulares sobre os mecanismos de resposta imune e sobre os processos que levam a quadros imunossupressivos e/ou anérgicos em patologias graves. (STEWART e NICHOLSON, 2000)
Para a determinação das linhagens celulares e para a análise da maturação das células nas neoplasias hematológicas, o método preferencialmente utilizado tem sido a caracterização imunofenotípica. O desenvolvimento da ampla gama de anticorpos monoclonais e das potencialidades do citômetro de fluxo têm impulsionado esta área nos últimos 20 anos. A citometria de fluxo é uma ferramenta metodológica que avalia as características fenotípicas, bioquímicas ou moleculares de células. O citômetro de fluxo pode avaliar propriedades como tamanho, granulosidade, além da complexidade interna, como componentes internos, externos e conteúdo de DNA, avaliados através de tipos de fluorescências obtidas do sangue periférico dos animais utilizando-se o software cell quest acoplado ao citômetro. (STEWART e NICHOLSON, 2000).
O objetivo da presente investigação foi avaliar alguns parâmetros fenotípicos antes e após a irradiação de células do sangue periférico (PBMC) de cobaias, através da técnica de citometria de fluxo, utilizando marcadores de linfócitos T e B, monócitos, estimulação e imunossupressão celular.
Materiais e métodos
O procedimento consistiu na obtenção de amostras de sangue periférico de cobaias para obtenção de perfil de expressão dos marcadores de ativação, imunossupressão, apoptose utilizando o citômetro de fluxo após a irradiação e contagem do número de linfócitos antes e após cinética de tempo. O projeto de pesquisa foi aprovado em 03 de fevereiro de 2006 pelo CPPPA- Centro de Preparações de Peças e Pesquisas Anatômicas da Faculdade de Ciências Médicas de Minas Gerais (FCMM), com autorização de uso do Biotério da FCMM, e utilização de cães no experimento, documentado e aprovado seguindo o regimento interno da faculdade, assinado pelo coordenador e professor titular Prof. Chrystiano Gonçalves Rocha. Levou-se em conta também a literatura sobre os aspectos éticos e morais da pesquisa em animais, preconizados por HOFF, 1980; FLECKNELL, 1993; FONSECA et al., 1996; PETROIANU, 1996 e 2000. Foram pré-estabelecida as doses de 2Gy, 4Gy e 6Gy. A cinética de tempo da coleta das amostras foi T0, antes da irradiação, grupo controle, T2 de 2h após a irradiação, T18 de 18h após a irradiação e T30, 30 dias após a irradiação. O procedimento envolveu irradiação de 2 Gy (n=2); 4 Gy (2 Gy fracionado, n=2); 6 Gy (2 Gy fracionado, n=1), empregando THERATRON 80 - cobalto 60, tamanho do campo: 13 x 17 cm2 - campo único – esquerdo, profundidade de tratamento: 2,5cm, sítio de irradiação da cabeça e pescoço com proteção de chumbo para os olhos, com doses fracionadas de 2Gy semanais. O lado esquerdo da região da cabeça e pescoço foi escolhido por atingir um território vascular bastante representativo.
As cobaias foram anestesiadas a cada irradiação com Dopalen® (quetamina), 5mg.kg-1 de peso e Rompum® (xylazina) na dose de 2mg.kg-1 de peso, por via parenteral intramuscular. A cobaia foi posicionada em decúbito dorsal na mesa operatória com imobilização das patas, com hiperextensão cervical e rotação lateral esquerda da cabeça. Procedida a depilação da região inguino-crural esquerda após palpação abdominal de rotina, fez-se a punção da artéria femoral direita, com agulha de 25 x 7mm montada em seringa descartável de 20 mL, sendo aspirado em torno de 8 mL a 10 mL de sangue. Após esse procedimento cada animal foi liberado para a recuperação anestésica em baia individual com água e ração sendo oferecidas de forma livre.
As células do sangue periférico-PBMC foram separadas e submetidas às marcações com os anticorpos monoclonais anti- CD5, anti-CD4, anti_IgM, anti-CD14, anti-MHCII, anti-CTLA-4 e anti-rIL-2 para avaliação das células T circulantes, células B, linhagem monocítica/macrofágica e parâmetros de estimulação, expansão clonal e regulação da expansão clonal, respectivamente.
As PBMCs foram separadas em tubos de 50 mL (Pyrex Laboratory Glassware®), siliconizados contendo uma mistura de Ficoll-diatrozato, obtida comercialmente (LSM® - Lymphocyte Separation Medium, Organon Teknika Corporation, Durhan, NC), na proporção de uma parte de Ficoll-diatrozato para duas partes de sangue.
A preparação foi submetida à centrifugação por 40min, a 400 X g à temperatura ambiente. Ao final da centrifugação obtivemos um anel de células mononucleares na interface entre o Ficoll e o plasma, o qual foi cuidadosamente removido com o auxílio de uma pipeta Pasteur (Thomas Laboratory Specialities®) e transferido para tubos de fundo cônico de 50ml estéreis (2070 Graduated conical tube, Falcon). O volume foi completado para 50ml com meio de cultura MEM (Minimal Essential Medium, GIBCO) e o frasco levado a centrifugação de 10min, 400 X g, 4C.
As células foram lavadas por três vezes utilizando-se cerca de 30ml de MEM. Ao final, as células foram ressuspendidas em 2ml de meio de cultura RPMI-1640 (GIBCO). Uma alíquota da suspensão de células foi diluída (1:20) em solução de Turck's e o número de células determinado através de contagem em câmara de Newbauer com o auxílio de um microscópio ótico. A concentração de células foi ajustada para uma suspensão contendo 10 x 106 células.ml-1 de RPMI-1640. Toda manipulação das células foi realizada em condições estéreis em capela de fluxo laminar (BBL®-Biological Cabinet, model 60474).
Os ensaios de imunofluorescência para população de células mononucleares de sangue periférico ex-vivo aconteceu segundo o protocolo indicado pela Becton Dickinson®, e adaptado para placas de 96 poços. Em cada poço foram adicionados 25ml de solução de anticorpo monoclonal específico para o receptor celular de interesse, marcados com fluorocromo, diluídos 1:5 em tampão FACS (PBS 1% BSA, pH 7.4). Para cada teste foram utilizados 25ml de células mononucleares, o equivalente a 250.000 células por poço. As placas foram incubadas por 20 a 30min ao abrigo da luz a 4°C. As amostras foram lavadas com 200ml de PBS 0.015M pH 7.4 e postas em centrifugação (1300rpm, 7min à 4°C) e o sobrenadante desprezado. Fixaram-se as amostras com 200ml de solução fixadora (10 g.l-1 de paraformaldeído, 1% de Cacodilato de Sódio, 6.67 g.l-1 de cloreto de Sódio, pH 7.2). Após um período de 30min a 4°C, as amostras foram transferidas para tubos de 1ml, e os parâmetros fenotípicos das células presentes em cada amostra foi determinado com o auxílio de um citômetro de fluxo (FACScan Becton Dickinson®). A identificação das populações celulares de interesse bem como a determinação do valor percentual de populações, subpopulações celulares e a expressão de moléculas co-estimuladoras nessas populações foram feitas utilizando-se o software cell quest acoplado ao citômetro.
O software estatístico utilizado foi teste-t de Student, e software ANOVA.
Resultados e discussão
Os achados em cinética de dose foram pouco expressivos sem diferenças estatísticas significantes. Então, optou se por analisar os dados pelas alterações fenotípicas ocorridas após a irradiação na cinética de tempo (T2, T18 e T30), seguindo como referência o padrão fenotípico registrado no tempo zero (T0), anterior à irradiação referente ao grupo controle. As populações celulares foram identificadas utilizando a análise dos histogramas de fluorescência. Foram analisadas em canal médio de fluorescência as células ou “eventos” que passam pelo feixe de laser e são marcadas com os anticorpos monoclonais anti- os receptores de interesse, com as fluorescências distintas. Esse tipo de análise nos permitiu obter o número absoluto de células e estimar a razão entre as populações marcadas com cada anticorpo.
Freqüência de linfócitos T CD5+ circulantes
A analise fenotipica dos linfócitos T foi realizada com o emprego dos anticorpos monoclonais anti-CD5. A Figura 1 mostra o percentual de linfócitos T CD5+ na população de linfócitos totais do sangue periférico antes e após a irradiação. O histograma de fluorescência teve a intensidade de fluorescência avaliados num canal médio de fluorescência de 0 a 1023, para o número absoluto de receptores para CD5, numa cinética de tempo no controle (zero), 2h, 18h e 30 dias após a irradiação com cobalto 60, considerando os valores positivos, a partir do canal 200 (a partir de M1). Foram encontradas diferenças estatisticamente significativas (p < 0,05) em relação ao tempo zero.
A análise dos resultados demonstrou diferença estatisticamente significativa antes e após a irradiação, correspondentes aos tempos avaliados para 2h, 18h e 30 dias, mas o mesmo não foi observado em relação as doses de 2, 4 e 6Gy. O percentual de linfócitos T CD5+ foi significativamente menor após as 18h após a irradiação quando comparada aos tempos de 2h e também no trigésimo dia pós-irradiação.
A função de CD5 nas células T não está muito bem estabelecida, mas tem sido reportado em humanos que esta atua como co-sinalizador direto para ativação ou co-estimulação, papel também exercido por CD2, CD4, CD6 e CD28. Esse receptor pode estar envolvido na mobilização de Ca2+ intracelular, além de contribuir para o aumento da reposta mitogênica de secreção e expressão de IL-2. (LANDMANN et al, 1998; ANTONY, 2005)

Freqüência de linfócitos B IgM+ circulantes
A população de linfócitos B foi identificada com o emprego dos anticorpos monoclonais anti-IgM canina. A Figura 2 mostra o percentual médio de linfócitos B IgM+ na população de linfócitos totais do sangue periférico antes e após a irradiação. O histograma de fluorescência foi analisado conforme os canais descritos anteriormente. Diferenças estatisticamente significativas (p < 0,05) em relação ao tempo zero foram observadas, antes e após a irradiação. O percentual de linfócitos B foi significativamente menor após 2h e 18h da irradiação sendo 27,12 ± 0,95% e 23,02 ± 0,65%, respectivamente, em relação ao controle de 34,11. ± 1,34% (p <0,05). Os dados mostraram, ainda, diminuição do percentual dessas células a 30 dias após a irradiação, de 25,21. ± 2,13%, quando comparados com o controle.
Sabe-se que a resposta tipo 2 gera aumento de IL4 que estimula a diferenciação de linfócitos B, ocasionando aumento na produção de anticorpos. Esse padrão encontra-se associado comumente a uma doença severa, que pode estar enquadrada como uma parasitose. No caso do nosso modelo, que avalia a imunossupressão induzida pela irradiação, a própria diminuição do padrão CD4 pode levar, concomitantemente, à diminuição dos linfócitos B diferenciados (plasmócitos) e, conseqüentemente, à diminuição da resposta IgM+.
REIS, 2001, explica a diminuição do percentual de células B considerando que, no processo de transformação dos linfócitos B em diferenciados, há alterações no tamanho e granulosidade dessas células, o que pode promover um deslocamento da população para fora da região de linfócitos analisados durante a citometria de fluxo.

Pode-se também levantar a possibilidade de reação cruzada oriunda de outras infecções permanentes ou transitórias no modelo animal que embora sem sintomas clínicos podem apresentá-las em período de incubação ou em estado sub-clínico. Assim, a não detecção de diferenças estatisticamente significativas no percentual de células B IgM+ poderia estar relacionada com uma possível ineficiência do modelo animal em experimentação. (UCHOA et al. 2001)
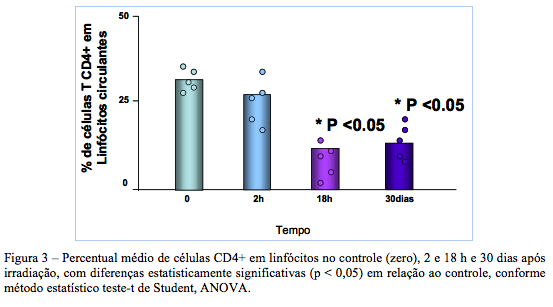
Freqüência de linfócitos T CD4+ circulantes
O percentual de células T CD4+ na população de linfócitos circulantes antes e após irradiação é apresentado na Fig. 3. A análise do histograma de fluorescência seguiu os mesmos procedimentos descritos anteriormente.
A analise estatística dos resultados demonstrou uma redução da população celular no tempo de 2h após a irradiação (26,67 ± 4,23%) em relação ao tempo zero (36,65 ± 2,56%). A redução do percentual foi significativa com p<0,05 nas 18h seguinte, onde se observou um percentual médio de células T circulantes de 12,45 ± 2,95%, que pode ser indicativo do efeito acumulativo da dose absorvida de irradiação gama, percentual esse que se manteve ainda bastante reduzido quando avaliado no trigésimo dia após irradiação.
No entanto, vale ressaltar que dados da literatura já demonstraram a expressão do marcador CD4 também por neutrófilos em modelo de infecção experimental por parasitas em cães no qual há estimulação e recrutamento de outras linhagens celulares, não sendo observada em outras espécies de mamíferos (MOORE et al, 1992),.
Os resultados apontaram a redução do marcador CD4 já nas primeiras 2h após a irradiação. Esta redução pode estar associada ao modelo de imunossupressão, no qual geralmente há a inversão do padrão CD4:CD8 que normalmente atinge 2:1.
Há indicativo que a redução observada neste modelo canino pode estar associada a uma provável imunosupressão pós-exposição, possivelmente presente em situações clínicas humanas na radioterapia. É interessante notar que em um modelo de vacinação com peptídeos específicos E5 observa-se exatamente o oposto visto que o peptídeo utilizado na vacinação foi capaz de induzir o recrutamento tanto de subpopulações CD4+ e CD8+ “naive” ou“virgens” ressaltando em uma resposta imune de memória estável. Em adição, a vacinação induziu a ativação geral de célula T total em específico para o efetor de CD4+. A vacinação com E5 levou à ativação de células T de memória CD45RO+, CD4+, CD8+ “naive” ou “virgens”; enquanto, paralelamente, recrutou subpopulações CD4+ e CD8+ da resposta imune geral. (HUEMAN, 2006)
Considerando que em nosso estudo observamos uma diminuição de células CD4+ após a irradiação, provavelmente poderá também haver uma diminuição da subpopulação CD4CD45RO+, uma vez que no trigésimo dia pós-irradiação a freqüência de células CD4 ainda permaneceu baixa em relação ao tempo zero no controle não irradiado.
Podemos inferir, ainda, que os efeitos observados nesse estudo podem estar associados à perda de ativação de enzimas essenciais que conferem a co-sinalização para expansão clonal, interferindo possivelmente na transcrição do IL-2R.
A redução da capacidade proliferativa pode estar associada a uma possível alteração da capacidade de reparo das células irradiadas, processo que normalmente se completa poucas horas depois da irradiação. Esta informação corrobora com a observação de uma elevação no percentual de células CTLA-4+ verificado após a irradiação.

Freqüência de linfócitos T CD5+ circulantes co-expressando o receptor CTLA-4
A análise fenotípica dos linfócitos T após a irradiação foi realizada com o emprego dos anticorpos monoclonais anti-CD5, co-expressando o receptor CTLA-4. CTLA-4 é um anticorpo quimérico usado em doenças autoimunes, como asma, infecções e rejeição a enxertos. Seu alvo consiste em bloquear a co-estimulação de linfócitos T através da inibição da ligação das moléculas B7. O bloqueio desta ligação está associado a anergia, apoptose e indução de tolerância em modelos experimentais de doenças autoimunes e transplante.
A Figura 4 evidencia o percentual de linfócitos T CD5+CTLA-4+ na população de linfócitos totais do sangue periférico antes e após a irradiação, com diferenças estatisticamente significativas, correspondentes aos tempos avaliados para 2h, 18h e 30 dias. A razão entre o percentual de linfócitos T CD5+ que co-expressaram o receptor CTLA-4 foi significativamente maior nas duas primeiras 2h após a irradiação, quando comparada com o controle, ocorrendo um pico às 18h.
Considerando que uma vez que as células T estejam ativadas, elas passam a exibir um receptor adicional, designado como CTLA-4, que se assemelha muito com a seqüência de CD28. Essa elevação do número de receptores para CTLA-4 pós-irradiação pode ser inicialmente justificado pela necessidade de regulação da expansão clonal. Sabe-se que as moléculas B7, presentes na superfície das APCs, ligam-se com maior avidez a CTLA-4 presente na superfície de células T, do que a CD28. A ligação de B7 a CTLA-4 é capaz de emitir um sinal inibitório à célula T ativada para que a resposta proliferativa seja efetivamente interrompida e para que seja produzida uma menor quantidade de IL-2, que é um fator de crescimento para a célula T. Assim, a ligação de CTLA-4 às moléculas B7 é essencial para diminuição da resposta proliferativa das células T ativadas pelo antígeno e à molécula B7 na superfície das APCs. Vale ressaltar que, a utilização do nosso modelo não nos possibilita caracterizar uma resposta antígeno-específica, visto que não há presença de um antígeno. Mas, podemos inferir que o estímulo exógeno da radiação confere o “start” para ativação celular e para regulação da resposta celular.
Freqüência de linfócitos T CD5+ circulantes co-expressando o receptor CTLA-4
A análise fenotípica dos linfócitos T após a irradiação foi realizada com o emprego dos anticorpos monoclonais anti-CD5, co-expressando o receptor CTLA-4. CTLA-4 é um anticorpo quimérico usado em doenças autoimunes, como asma, infecções e rejeição a enxertos. Seu alvo consiste em bloquear a co-estimulação de linfócitos T através da inibição da ligação das moléculas B7. O bloqueio desta ligação está associado a anergia, apoptose e indução de tolerância em modelos experimentais de doenças autoimunes e transplante.
A Figura 4 evidencia o percentual de linfócitos T CD5+CTLA-4+ na população de linfócitos totais do sangue periférico antes e após a irradiação, com diferenças estatisticamente significativas, correspondentes aos tempos avaliados para 2h, 18h e 30 dias. A razão entre o percentual de linfócitos T CD5+ que co-expressaram o receptor CTLA-4 foi significativamente maior nas duas primeiras 2h após a irradiação, quando comparada com o controle, ocorrendo um pico às 18h.
Considerando que uma vez que as células T estejam ativadas, elas passam a exibir um receptor adicional, designado como CTLA-4, que se assemelha muito com a seqüência de CD28. Essa elevação do número de receptores para CTLA-4 pós-irradiação pode ser inicialmente justificado pela necessidade de regulação da expansão clonal. Sabe-se que as moléculas B7, presentes na superfície das APCs, ligam-se com maior avidez a CTLA-4 presente na superfície de células T, do que a CD28. A ligação de B7 a CTLA-4 é capaz de emitir um sinal inibitório à célula T ativada para que a resposta proliferativa seja efetivamente interrompida e para que seja produzida uma menor quantidade de IL-2, que é um fator de crescimento para a célula T. Assim, a ligação de CTLA-4 às moléculas B7 é essencial para diminuição da resposta proliferativa das células T ativadas pelo antígeno e à molécula B7 na superfície das APCs. Vale ressaltar que, a utilização do nosso modelo não nos possibilita caracterizar uma resposta antígeno-específica, visto que não há presença de um antígeno. Mas, podemos inferir que o estímulo exógeno da radiação confere o “start” para ativação celular e para regulação da resposta celular.

Os perfis de células T totais e de suas subpopulações sugerem uma inversão do padrão CD4:CD8 após irradiação. Assim, esses dados corroboram com a literatura, uma vez que KUHN at al.,2000, mostraram que justamente a subpopulação CD8+CTLA-4+ foi capaz de regular a reatividade e a função das células CD4, o que justifica essa diminuição após a irradiação. Esses achados foram mais tarde confirmados por PARRY at al., 2005, que também demonstraram o papel regulador de CTLA-4, uma vez que esse receptor foi capaz de inibir a estimulação celular; e, consequentemente, o reconhecimento de peptídeos via CD3CD28 por outras populações CD86+ (B-7) do sistema imune.
Essas características podem ser observadas também em respostas anti-tumorais, quando HODI at al., (2003) apresentou que o receptor para CTLA-4 foi capaz de bloquear a imunidade tumoral em pacientes com carcinoma ovariano, quando foram previamente vacinados aumentando a apresentação dos produtos derivados do tumor por células dendríticas e limitando assim a potência da imunidade tumoral.
QUEZADA at al.,2006), demonstraram algo semelhante com pacientes com encéfalomielite, sendo observado uma diminuição de células CD4+ quando células CD8CTLA-4 eram estimuladas.
Freqüência de linfócitos T CD5+ circulantes co-expressando o receptor IL-2R
A análise dos parâmetros fenotípicos dos linfócitos T após a irradiação foi realizada com a marcação dos anticorpos monoclonais anti-CD5, co-expressando o IL-2R. A Figura 5 mostra o percentual de linfócitos T CD5+IL-2+ na população de linfócitos totais do sangue periférico antes e após a irradiação. A análise dos dados demonstrou diferença estatisticamente significativa antes e após a irradiação, correspondentes aos tempos avaliados para 2h e 18h (13,11 ± 1,95%) e (5,83 ± 2,96%), respectivamente, e aos 30 dias. A razão entre o percentual de linfócitos T CD5+ que co-expressaram o IL-2R não foi significativamente menor nas duas primeiras 2h após a irradiação, quando comparada ao controle (0 h: 11,23 ± 3,05%), porém, observou-se uma drástica diminuição nas 18h após a irradiação com p <0.05. Esse achado pode estar associado ao papel autócrino exercido pelo receptor CTLA-4, bem como sobre outras células que proliferaram devido à co-sinalização e que, provavelmente, podem ter deixado de expandir-se clonalmente, devido ao papel de CTLA-4. No entanto, observou-se que os percentuais aos 30 dias com 12,23 ± 2,79% não se diferenciaram em relação ao controle, sugerindo que a ação de CTLA-4 é pontual na auto-regulação, mas pode não estar envolvida no processo de imunosupressão a médio ou longo prazo.
Os estudos selecionados são concordantes com os presentes achados, uma vez que a diminuição do IL-2R pode ser pontuada após a irradiação pelo aumento de células T CTLA-4+. Então, é correto supor que a resposta de IL-2R observada nas 2h após a irradiação, onde não houve uma diminuição importante, sugere um efeito residual da ação de IL-2R, visto que as células parecem conseguir manter sua capacidade proliferativa, bem como a sobrevivência e a respectiva ativação logo após a irradiação. Esse evento foi observado por PING JIN et al., 2006, que utilizaram IL-2R recombinante em culturas de 47 doadores de perfis imunológicos diferentes e obtiveram a mesma capacidade de ativação celular, mantendo os efeitos anti-apoptóticos.

Freqüência de monócitos circulantes na população de leucócitos totais
Na Figura 6 estão representados os percentuais de monócitos CD14+ na população de leucócitos totais antes e após a irradiação nas mesmas doses já descritas. A Figura 6 mostra o percentual médio de monócitos co-expressando o receptor para CD14+ no controle T0, e nos tempos 2,1h e 30d após irradiação. Diferenças estatisticamente significativas (p < 0,05) em relação ao controle foram observadas, pelo método teste-t Student, ANOVA, após a irradiação, correspondentes à 2h e 18h com percentuais médios de 1,12 ± 0,95% e 1,83 ± 0,56%, respectivamente. A análise dos resultados revelou uma diferença estatisticamente significativa (p < 0,05) quando comparados ao controle de 5,78 ± 0,74%, mostrando uma queda acentuada nas 2h e 18h após irradiação mantendo em 30 dias após irradiação a 1,98 ± 0,65%.
Os mecanismos pelos quais a glicoproteína CD14 é capaz de exercer sua função, independente de um fator mitogênico inespecífico do tipo lipopolissacáride-LPS não estão bem estabelecidos ainda, mas sabe-se que é capaz de agir como receptor para LPS ou como uma molécula solúvel no soro, independente de estar ancorada numa membrana celular mieloide. (LANDMANN et al, 1998) No caso específico do nosso estudo, onde foram avaliados o fenótipo de PBMCs no modelo canino, há de se supor que a marcação com anticorpo monoclonal anti-CD14 poderia ser então positiva para o canal médio de fluorescência antes da irradiação, já que tal expressão independeria de estimulação prévia. É importante ressaltar, no entanto, que em modelos de infecção por parasitas, por exemplo, a tendência seria de elevação dessa marcação já nas primeiras horas pós-estimulação, visto que a linhagem monocítica é uma das primeiras a serem recrutadas na resposta imune (MOREIRA, 2005). Porém, considerando que o modelo utilizado avaliou os aspectos de imunussupressão através de cinética de irradiação, a queda dos percentuais observados a partir das 2h parece ser plausível.

Freqüência de linfócitos T circulantes co-expressando MHCII
MHCII são proteínas heterodímeras compostas por duas cadeias polipeptídicas alfa e beta contribuindo com um domínio ao sítio de ligação à peptídeo e um domínio de suporte semelhante à imunoglobulina. MHCII são sintetizadas no retículo endoplasmático da célula, se transferem para o aparelho de golgi ligadas a uma terceira proteína, chamada cadeia invariante, que evita qualquer ligação prematura de peptídeos endógenos ao seu sítio de ligação, até que elas atinjam o local de degradação protéica extracelular. Uma segunda função da cadeia invariante é enviar as moléculas MHCII para as vesículas endocíticas onde elas se ligam à peptídeos.
A análise fenotípica dos linfócitos T após a irradiação foi realizada com o emprego dos anticorpos co-expressando MHCII, considerando que se trata de uma molécula que se liga aos peptídeos antigênicos para posterior apresentação às células T, e serve também como um sinal de tradução que regula a função de macrófagos (MATSUYAMA et al, 1993).
A Figura 7 mostra a expressão de MHCII+ no sangue periférico antes e após a irradiação. A análise estatística dos dados demonstrou não haver diferença estatisticamente significativa antes e após a irradiação, correspondentes aos tempos de 2 e 18h. A expressão da molécula MHCII foi menor nas duas primeiras horas após a irradiação quando comparada ao controle, sendo essa diminuição não estatisticamente significativa e nem tampouco às 18h com as doses correspondentes. Considerando que, em nosso estudo, a dupla marcação não foi efetuada juntamente com macrófagos, não foi possível avaliar se essa redução reflete o efeito da irradiação sobre a ativação de proteínas, tais como a tirosina Kinase, proteína Kinase C, já que essas vias de sinalização afetam a apresentação de antígenos.
É importante reconhecer também que, contrariamente aos achados de modelos de infecção por parasitas (CASTRO, 2004), em nosso estudo não houve sinalização para apresentação de antígenos e as modificações celulares ocorridas podem ter inviabilizado a expressão de MHCII+ devido à irradiação agindo assim como fator inibitório. Entretanto, estudos mais aprofundados são necessários.

De qualquer forma, sabe-se que a ligação do complexo peptídeo MHC ao receptor de célula T (TCR) e a ligação de B7 a CD28 na célula T atuam da seguinte forma: i) dirigem a ativação e expansão de clones de células T, que encontram o antígeno específico apresentado por uma APC; ii) dirigem a ativação das células T “virgens”, que sejam capazes de induzir a expressão e a secreção de interleucina 2 (IL-2); iii) ampliam a expressão de receptores de alta afinidade para IL-2 na superfície das células T antígeno-específicas. Então torna-se coerente inferirmos sobre uma possível relação entre a diminuição da expressão do receptor para MHC, já nas primeiras horas pós-irradiação, e a uma falha na ativação das vias enzimáticas responsáveis pela tradução do mensageiro em proteína pela própria irradiação.
Sabe–se que a radiação ionizante pode produzir mudanças estruturais em várias classes de macromoléculas encontradas nas células. A radiação resulta na indução de genes de respostas aguda e tardia, sendo que o produto desses genes, especialmente as citocinas e fatores de crescimento, podem influenciar a resposta histoquímica dos tecidos (ROSENTHAL et al, 1995).
Dados experimentais, providos por EISBRUCH et al, 1999, também sugerem que os fatores de crescimento são sintetizados e secretados pelas células sobreviventes à radiação. Moléculas semelhantes ao fator de crescimento secretado da parede dos vasos sanguíneos podem promover proliferação de células musculares lisas, participar da proliferação anormal de células endoteliais, e promover o início da fibrose pós-irradiação. A proliferação de células endoteliais e das células do músculo liso em artérias de pequeno calibre, seguida pelo estreitamento na luz dos vasos sanguíneos, tem sido observada após a irradiação produzindo um decréscimo no fluxo sanguíneo, levando a anóxia, necrose e substituição dos elementos constituintes do tecido normal por fibrose. (SALVAJOLI et al, 1999).
Conclusão
Esse estudo demonstrou o comportamento do fenotípico de linhagens PBMC alteradas pela irradiação em modelo animal. Podemos inferir sobre a importância do conhecimento da resposta imune individualizada, através da investigação do padrão fenotípico das linhagens celulares periféricas circulantes sistêmicas, para facilitar o entendimento da imunosupressão.
Acreditamos que a continuidade de tal estudo através de ensaios clínicos possibilitará ampliar a qualidade de vida de pacientes submetidos à radioterapia externa, visando monitorar e melhorar a resposta do sistema imunológico do paciente e consequentemente atingir sucesso no tratamento do câncer.
Referências
ANTONY P. A., PICCIRILLO C. A. J Immunol. Março; 174(5): páginas 2591–2601, 2005.
CASTRO, M. B.. Doutorado. Caracterização imunoistoquímica das células mononucleares em lesões observadas na infecção aguda experimental em cães por Erlichias canis. Faculdade de Ciências Agrárias e Veterinárias de Jaboticabal, Unesp, Jaboticabal, página 76, 2004.
DEVITA, V.T.; HELLMAN, S.; ROSENBERG, S.A. Cancer : principles and practice of oncology. Philadelphia: J. B. Lippincott, página 1926, 1982.
FLECKNELL, P.A. British Journal of anaesthesia. Anaesthesia animals for biomedical research. Vol.71, p 885 a 894, 1993.
FONSECA, M.N., GOLDENBERG, S., GOMES, P.O. Anestesia em coelhos. Acta Cir. Bras. v. 11, páginas 82-104, 1996.
FRANKS, L.M. Introdução à biologia celular e molecular do cancer. São Paulo: Roca, 1990. 423 p.
GREENWALD, R.J et al. B7-2 is required for the progression but not the initiation of the type 2 immune response to a gastrointestinal nematode parasite. J. Immunol., Baltimore, v. 162, p. 4133 – 4139, Apr. 1999.
HOFF, C. Immoral and moral uses of animals. N. Eng. J. Med., v. 302, p. 115-8, 1980.
HUEMAN M.T., STOJADINOVIC A., et al. Analysis of naive and memory CD4 and CD8 T cell populations in breast cancer patients receiving a HER2/neu peptide (E75) and GM-CSF vaccine. PMID: 16783576 PubMed as supplied by publisher, 2006.
KUHNS M.S., EPSHTEYN V., et al. Cytotoxic T lymphocyte antigen-4 (CTLA-4) regulates the size, reactivity, and function of a primed pool of CD41 T cells. PNAS. vol. 97 no. 23 nov 7, 2000.
LANDMANN R., LINK S., et al. Soluble CD14 Activates Monocytic Cells Independently of Lipopolysaccharide. Infect Immun. 1998 Maio; 66(5): 2264–2271.
MATSUYAMA, S., KOIDE, Y., YOSHIDA, T.O. HLA class II molecule-mediated signal transduction mechanism responsible for the expression of interleukin-1 beta and tumor necrosis factor-alpha genes induced by a staphylococal superantigen. Eur. J. Immunol. n.23, p.3194-3202, 1993.
MOORE, P. F.; ROSSITO, P. V.; DANILENKO, D. M. et al. Monoclonal antibodies specific for canine CD4 and CD8 define functional T- lymphocyte subsets and high-density expression of CD4 by canine neutrophils. Tissue Antigens, n. 40, p. 75-85, 1992.
PARRY R.V., CHEMNITZ J.M., et al. CTLA-4 and PD-1 Receptors Inhibit T-Cell Activation by Distinct Mechanisms. Molecular and cellular biology. American Society for Microbiology. p. 9543–9553 vol. 25, no.21, nov, 2005.
PETROIANU, A. In. PETROIANU, A. Pesquisa experimental. Ética, Moral e Deontologia Médicas. Rio de Janeiro 1ª. ed., Guanabara-Koogan, p. 90 -185, 2000.
PETROIANU, A. Aspectos éticos na pesquisa em animais. Acta Cir. Bras., v.11, p. 64-157, 1996.
PEREZ, C.A.; TAVEIRA, L.A.. Principles and practice of radiation oncology. 3. ed. Philadelphia: Lippincott-Raven, 1998.
PING J., WANG E., et al. Molecular signatures induced by interleukin-2 on peripheral blood mononuclear cells and T cell subsets. J Transl Med. Junho, 2006.
QUEZADA S.A., PEGGS K.S. et al. CTLA4 blockade and GM-CSF combination immunotherapy alters the intratumor balance of effector and regulatory T cells. Howard Hughes Medical Institute, Department of Immunology, Memorial Sloan-Kettering Cancer Center. The Journal of Clinical Investigation. New York, USA. v. 116 n. 7, julho, 2006.
REIS, A.B. Avaliação de parâmetros Laboratoriais e Imunológicos de Cães naturalmente Infectados com Leishmania (Leishmania) chagasi portadores de Diferentes Formas Clínicas da Infecção. Tese de Doutorado, Instituto de Ciências Biológicas, 115p, 2001.
ROSENTHAL, S.N. Oncologia prática: cuidados com o paciente. 2. ed. Rio de Janeiro: Revinter, p339,1995.
SALVAJOLI, J.V.; FARIA, S.L.. Radioterapia em oncologia. Rio de Janeiro: MEDSI, p1243, 1999.
STEWART, C.C.; NICHOLSON, J.K.A. Immunophenotyping. New York: Wiley-Liss, p448, 2000.
UCHOA, C.M.A.; SERRA, C.M.B, et al.. Aspectos sorológicos e epidemiológicos da leishmaniose tegumentar americana canina. Marica. RJ, Brasil. Rev. Soc. Bras. Méd. Trop. v.34 n.6. Uberaba nov./dic., 2001.